CRESCIMENTO
INFANTIL-JUVENIL: OS FATORES DE CRESCIMENTO SEMELHANTES À INSULINA-IGFS CIRCULAM
LIGADOS A PROTEÍNAS DE LIGAÇÃO DO FATOR DE CRESCIMENTO -IGFBPS, SEIS DOS QUAIS
FORAM IDENTIFICADAS. A IGFBP-3 É A PRINCIPAL IGFBP CIRCULANTE DURANTE A VIDA
PÓS-NATAL E É GH-DEPENDENTE


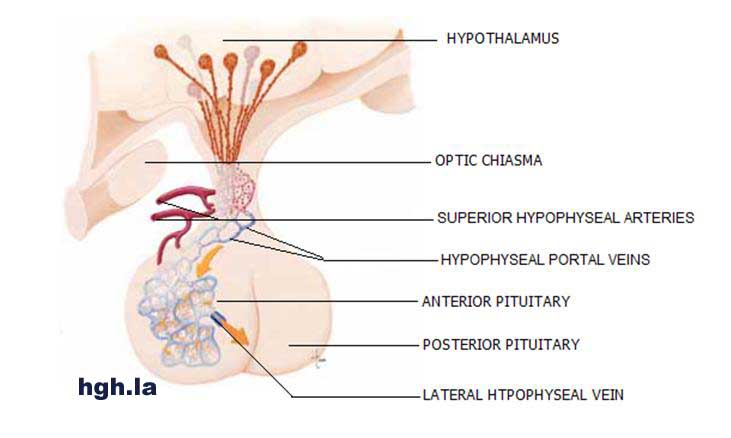
Os aumentos foram amplamente
informados em concentrações de GH basal, a frequência de pulso e amplitude de
GH em pacientes com diabetes têm profundos efeitos anti-insulina semelhante à
de indivíduos saudáveis, resultando em aumento. As concentrações de glicose no
sangue associadas com a diminuição da sensibilidade periférica à insulina,
apesar de níveis mais altos de secreção de insulina, o aumento da produção de glicose
hepática, a estimulação da lipólise e a diminuição de insulina-estimulada pela
captação de glicose no músculo. Em indivíduos saudáveis o GH exógeno
administrado em níveis fisiológicos provoca resistência à insulina, por
prejudicar a capacidade da insulina para suprimir a utilização da glicose
hepática. Este efeito é caracterizado por reduzida afinidade hiperglicêmica de
ligação ao receptor da insulina, assim como um provável defeito de
pós-receptor. Assim, o desenvolvimento da resistência à insulina em indivíduos
com a função das células beta normal está associado com um aumento
compensatório na secreção de insulina, que permite a continuação da manutenção
da tolerância à glicose normal. Em indivíduos com DM1, efeitos semelhantes
foram demonstrados pelo aumento da necessidade de insulina durante os estudos.
 De fato, usando uma taxa variável de insulina durante a noite em infusão de
protocolos de manutenção euglicêmicos, alguns autores têm demonstrado uma
relação positiva entre sensibilidade periférica à insulina e as concentrações
de GH durante a noite atribuível ao efeito do GH sobre o metabolismo da glicose
periférica. De fato, uma correlação linear positiva a variações nas
concentrações séricas de GH tem sido descrita. Os níveis aumentados de GH resultam
em hiperglicemia septicemia hemorrágica viral que conduz ao aumento da noite
para o dia das necessidades de insulina característicos do “fenômeno dawn”.
Embora muitos estudos têm sido realizados para esclarecer as ligações entre GH
e a hipersecreção e sensibilidade periférica à insulina em DM1, o mecanismo
molecular ainda não foi esclarecido.
De fato, usando uma taxa variável de insulina durante a noite em infusão de
protocolos de manutenção euglicêmicos, alguns autores têm demonstrado uma
relação positiva entre sensibilidade periférica à insulina e as concentrações
de GH durante a noite atribuível ao efeito do GH sobre o metabolismo da glicose
periférica. De fato, uma correlação linear positiva a variações nas
concentrações séricas de GH tem sido descrita. Os níveis aumentados de GH resultam
em hiperglicemia septicemia hemorrágica viral que conduz ao aumento da noite
para o dia das necessidades de insulina característicos do “fenômeno dawn”.
Embora muitos estudos têm sido realizados para esclarecer as ligações entre GH
e a hipersecreção e sensibilidade periférica à insulina em DM1, o mecanismo
molecular ainda não foi esclarecido.

{kind=link}
Portanto, vários estudos com GH DNA-recombinante exógeno humano (rh)
IGF-I, em associação com a insulinoterapia foram realizados. Exógena rhIGF-1.
Como saber mais:
1. As ações e interações dos sistemas endócrino e nervoso, por meio das quais o sistema nervoso regula o sistema endócrino e a atividade endócrina modula a atividade do sistema nervoso central, constituem os principais mecanismos reguladores de praticamente todas as atividades fisiológicas...
http://hormoniocrescimentoadultos.blogspot.com.
2. Células nervosas e células endócrinas, ambas envolvidas na comunicação entre as células, compartilham certas características-secreção de mensageiros químicos (neurotransmissores ou hormônios) e atividades elétricas...
http://longevidadefutura.blogspot.com
3. Um mensageiro químico isolado-peptídeo ou amina-pode ser secretado por neurônios como um neurotransmissor ou por hormônio neural e por células endócrinas como um hormônio clássico...
http://imcobesidade.blogspot.com
AUTORIZADO O USO DOS DIREITOS AUTORAIS COM CITAÇÃO
DOS AUTORES PROSPECTIVOS ET REFERÊNCIA BIBLIOGRÁFICA.
Referências Bibliográficas:
Dr. João Santos Caio Jr, Endocrinologista, Neuroendocrinologista, Dra Henriqueta Verlangieri Caio, Endocrinologista, medicina interna – Van Der Häägen Brazil, São Paulo, Brasil; Salardi S, S Tonioli, Tassoni P, Tellarini M, L & Mazzanti Cacciari E. Crescimento e fatores de crescimento em diabetes Mellitus. Archives of Doenças na Infância 1987 62 57-62;
Penfold J, Chase HP, Marshall G, Walravens CF, Walravens PA & Garg SK.
Altura adulta final e sua relação com a glicose no sangue controle e
complicações microvasculares em IDDM. Diabetic Medicine 1995 12 129-133;
Donaghue KC, Kordonouri O, Chan A & M. Silink Secular tendências no
crescimento em diabetes: estamos ganhando? Arquivos de Doenças na Infância 2003
88 151-154; Kember NF. Cinética das células e o controlo do crescimento de
ossos longos. Cinética tecido celular 1978 11 477-485; Kember NF. Cinética das
células e o controlo do crescimento do osso. Acta Paediatrica 1993 82 (Suppl
391) 61-65; Salmão WD Jr & Daughaday WH. Um soro hormonalmente controlado
fator que estimula a incorporação de sulfato de cartilagem in vitro. Jornal de
Laboratório e Clínica 1957 49 825-836; Salmon WD Jr & Daughaday WH. Um soro
hormonalmente controlado fator que estimula a incorporação de sulfato de
cartilagem in vitro. Jornal de Laboratório e Clínica 1990 116 408-419; Jones JI
& Clemmons DR. Fatores de crescimento semelhante à insulina e sua proteínas
de ligação: ações biológicas. Endocrine Comentários 1995 163-34; Blum W &
Ranke MB. Fator de crescimento semelhante à insulina proteínas de ligação
(IGFBPs), com especial referência a IGFBP-3. Acta Paediatrica Scandinavica 1990
(Suppl 367) 55-62; Francesco Chiarelli, Cosimo Giannini e Angelika Mohn
Departamento de Pediatria da Universidade de Chieti, Itália.
1987 62 57-62;
Penfold J, Chase HP, Marshall G, Walravens CF, Walravens PA & Garg SK.
Altura adulta final e sua relação com a glicose no sangue controle e
complicações microvasculares em IDDM. Diabetic Medicine 1995 12 129-133;
Donaghue KC, Kordonouri O, Chan A & M. Silink Secular tendências no
crescimento em diabetes: estamos ganhando? Arquivos de Doenças na Infância 2003
88 151-154; Kember NF. Cinética das células e o controlo do crescimento de
ossos longos. Cinética tecido celular 1978 11 477-485; Kember NF. Cinética das
células e o controlo do crescimento do osso. Acta Paediatrica 1993 82 (Suppl
391) 61-65; Salmão WD Jr & Daughaday WH. Um soro hormonalmente controlado
fator que estimula a incorporação de sulfato de cartilagem in vitro. Jornal de
Laboratório e Clínica 1957 49 825-836; Salmon WD Jr & Daughaday WH. Um soro
hormonalmente controlado fator que estimula a incorporação de sulfato de
cartilagem in vitro. Jornal de Laboratório e Clínica 1990 116 408-419; Jones JI
& Clemmons DR. Fatores de crescimento semelhante à insulina e sua proteínas
de ligação: ações biológicas. Endocrine Comentários 1995 163-34; Blum W &
Ranke MB. Fator de crescimento semelhante à insulina proteínas de ligação
(IGFBPs), com especial referência a IGFBP-3. Acta Paediatrica Scandinavica 1990
(Suppl 367) 55-62; Francesco Chiarelli, Cosimo Giannini e Angelika Mohn
Departamento de Pediatria da Universidade de Chieti, Itália.
Dr. João Santos Caio Jr.
Endocrinologia – Neuroendocrinologista
CRM 20611
Dra. Henriqueta V. Caio
Endocrinologista – Medicina Interna
CRM 28930
Como saber mais:
1. As ações e interações dos sistemas endócrino e nervoso, por meio das quais o sistema nervoso regula o sistema endócrino e a atividade endócrina modula a atividade do sistema nervoso central, constituem os principais mecanismos reguladores de praticamente todas as atividades fisiológicas...
http://hormoniocrescimentoadultos.blogspot.com.
2. Células nervosas e células endócrinas, ambas envolvidas na comunicação entre as células, compartilham certas características-secreção de mensageiros químicos (neurotransmissores ou hormônios) e atividades elétricas...
http://longevidadefutura.blogspot.com
3. Um mensageiro químico isolado-peptídeo ou amina-pode ser secretado por neurônios como um neurotransmissor ou por hormônio neural e por células endócrinas como um hormônio clássico...
http://imcobesidade.blogspot.com
AUTORIZADO O USO DOS DIREITOS AUTORAIS COM CITAÇÃO
DOS AUTORES PROSPECTIVOS ET REFERÊNCIA BIBLIOGRÁFICA.
Referências Bibliográficas:
Dr. João Santos Caio Jr, Endocrinologista, Neuroendocrinologista, Dra Henriqueta Verlangieri Caio, Endocrinologista, medicina interna – Van Der Häägen Brazil, São Paulo, Brasil; Salardi S, S Tonioli, Tassoni P, Tellarini M, L & Mazzanti Cacciari E. Crescimento e fatores de crescimento em diabetes Mellitus. Archives of Doenças na Infância
Site Van Der Häägen Brazil
www.vanderhaagenbrazil.com.br http://drcaiojr.site.med.br
http://dracaio.site.med.br
Video
http://youtu.be/woonaiFJQwY
CONTATO:
Fones: 55(11) 5572-4848 (11)2371-3337 - (11)9.8197-4706 - TIM
Fones: 55(11) 5572-4848 (11)2371-3337 - (11)9.8197-4706 - TIM
Rua Estela, 515 - Bloco D - 12º andar - Conj 121/122
Paraiso - São Paulo - SP - Cep 04011-002
e-mail: vandergaagenbrasil@gmail.com
Paraiso - São Paulo - SP - Cep 04011-002
e-mail: vandergaagenbrasil@gmail.com